人口质量与健康是全球关注的核心科学与社会问题。近年来,全球不孕不育问题不断加剧。据世界卫生组织数据显示,全球约1/6成人受到不孕不育困扰。伴随着环境变化和育龄推迟,新生儿遗传缺陷发生率逐渐上升。其中,染色体异常在新生儿中的比例高达1/150至1/120,而胚胎染色体数目异常的根源多源于减数分裂的异常。在此过程中,交叉重组的数目和排布异常导致生殖细胞染色体不稳定与新生儿遗传缺陷。
早在上世纪初, Sturtevant和Muller就发现减数分裂中的“交叉重组”在染色体上的非随机分布现象(doi:10.1007/BF01792906; doi:10.1086/279534):当一对染色体上出现多个交叉重组位点时,它们会相距较远,表现出相互干扰现象 (crossover interference)。虽然这一现象极其重要,人们却一直未能阐释其背后机制。
2025年6月 19日中国科学技术大学细胞动力学教育部重点实验室访问教授张良余博士、微尺度物质科学国家研究中心姚雪彪教授联合国际合作团队,在国际知名期刊《Nature Cell Biology》杂志上发表了题为 “Crossover patterning through condensation and coarsening of pro-crossover factors” 的研究长文。这项研究首次融合大量标准化实验数据与数理模型,系统揭示了减数分裂交叉重组在染色体上排布阵势的形成机理,在解答这一百年谜题上取得重要进展。

审稿人高度评价了该项工作: “The molecular mechanism on how crossover formation is regulated remains unresolved and under long debate in the field since the original discovery of crossover interference in the fruit fly by Dr. Sturtevant in 1913.” “This model explains crossover interference because the formation of a single droplet depletes ZHP-3 from the rest of the chromosome pair. This is a very strong contribution to an exciting field and should be published in Nature Cell Biology.”

在减数分裂过程中,每一对染色体都会程序性发生多处双链DNA断裂,但只有极少数断裂会被修复为交叉重组。促成这一过程的关键蛋白—交叉重组蛋白(pro-crossover factors)—在重组位点组装形成蛋白复合体,即重组结节(recombination nodule)。自上世纪70年代在果蝇中首次观察到重组结节以来,通过电子显微镜技术,科学家们发现其表现为椭球形的电子致密体,位于联会复合体上(synaptonemal complex, SC)。SC是一种通过液-液相分离,在配对的同源染色体间组装的生物大分子凝聚体。在大多数真核生物中,交叉重组的干扰现象依赖于联会复合体存在。减数分裂早期,染色体上含有大量早期重组结节,位于双链断裂位点,分布较为随机;随着细胞周期推进,重组结节数目逐渐减少,体积增大,形成成熟的晚期重组结节。这些晚期重组结节的分布即反映了交叉重组的空间排布规律(图1)。
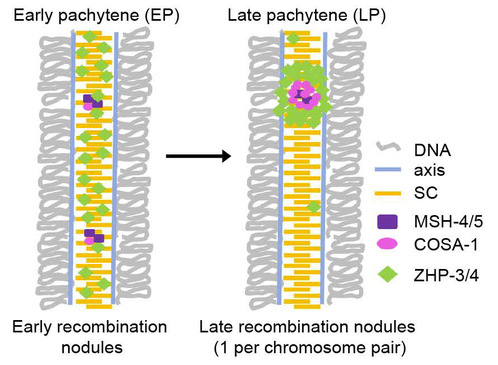
图1 联会复合体与重组结节示意图
联合团队选用秀丽隐杆线虫(C. elegans)为模式系统。线虫通体透明、繁殖周期短、遗传编辑易行,是该研究的理想模式体系。然而线虫系统一直缺乏高时空分辨率的条件性蛋白质敲除技术,为此,张良余博士在线虫中建立了植物生长素诱导的蛋白质条件性敲除系统(doi: 10.1242/dev.129635) 。这一技术使得研究必需基因在交叉重组过程中的时空动力学功能成为可能, 为解析交叉重组的发生和排布机制提供了关键基础。
在此工作中,研究团队采用荧光相关光谱技术和荧光漂白恢复等多种动态蛋白质定量分析手段,成功在活体动物生殖腺中表征了联会复合体和重组结节组分的动态性。结果显示,联会复合体和重组结节中的蛋白组分具有高度动态性,而每一条联会复合体作为相对独立的“区室”,在交叉重组的时间窗口内,限制关键交叉重组蛋白ZHP-3/4与核质的交换。同时,这些区室作为“基质”,使得ZHP-3/4沿着配对的染色体一维运动,从而协调同一对染色体上不同断裂位点的修复命运。更进一步,团队通过操控染色体断裂位点的数目以及染色体融合等手段,验证了联会复合体作为“区室单位”,控制着交叉重组蛋白的运动和每对染色体上交叉位点的生成和数量。细胞和体外实验还揭示了ZHP-3/4通过相分离形成蛋白凝聚体。借助遗传和细胞学分析,团队鉴定出一个位于重组结节的非经典蛋白激酶复合物CDK-2/COSA-1,并发现了一部分位于重组结节上的底物,揭示锚定于重组结节的蛋白激酶通过介导磷酸化,促进交叉重组蛋白在重组位点凝聚,从而形成类似活性液滴样结构(图2a)。结合上述的动态性分析和数学建模,研究团队发现早期重组结节中动态组分ZHP-3/4沿着每条联会复合体,通过奥斯特瓦尔德熟化的粗粒化过程,可以逐渐形成相互间隔较远的晚期重组结节(图2b)。该模型可以解释从低等真核生物酵母到人不同物种中所观察到的交叉重组排列阵势的形成过程。
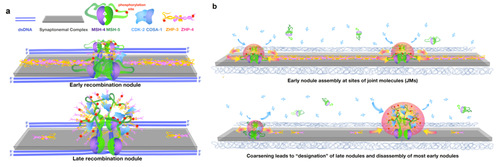
图2 重组结节的形成和粗粒化模型
该项工作不仅在揭示交叉重组干扰现象的形成机制这一百年谜题上取得重要突破,也发现了生物大分子凝聚态的新功能,即作为信号转导的基质,介导亚细胞水平分布阵势的形成,为解码生命与物质科学的共性科学问题提供了新的模式,并开拓了新方向。鉴于交叉重组决定减数分裂染色体的精确分离和健康生殖细胞的形成,本研究对人类不孕不育疾病的诊治具有重要的理论指导意义。
细胞动力学教育部重点实验室张良余访问教授、微尺度物质科学国家研究中心姚雪彪教授等为本文共同通讯作者;张良余博士同时为本工作的共同第一作者。本项研究得到了中国科大刘行教授以及多位国际合作者的宝贵帮助。
该研究得到国家自然科学基金委和科技部等项目支持。
原文链接:https://www.nature.com/articles/s41556-025-01688-9

细胞动力学教育部重点实验室 姚雪彪团队 部分成员合影